Far-Red Light and Phytochrome in LED Grow lights for Horticulture
- Apr 21
- 13 min read
Updated: Apr 22
From spectral theory to practical lighting strategies in controlled environment agriculture

Most discussions around horticulture lighting still revolve around the familiar red/blue ratio. It is a logical starting point: chlorophyll absorption peaks in those regions, and the quantum efficiency of Photosynthesis is highest in red light.
But for growers operating at the level of precision agriculture—where morphology, flowering timing, and biomass partitioning are actively engineered—this framing is incomplete.
Plants are not only responding to energy. They are interpreting signals.
And one of the most information-dense signals they receive is the balance between red and far-red light—processed through the phytochrome system.
The Phytochrome System: a continuously tuned biological switch

The phytochrome system operates as a dynamic light sensor that converts spectral information into developmental decisions. It is built around two interconvertible molecular states:
* Pr, which absorbs red light (~660 nm)
* Pfr, the biologically active form, which absorbs far-red light (~730 nm)
When red photons dominate, the system shifts toward Pfr. When far-red dominates, it shifts back toward Pr. This conversion is rapid, reversible, and constantly responsive to the spectral environment.
What matters in practice is not the absolute amount of red or far-red light, but their ratio—captured in a parameter known as the Phytochrome Stationary State (PSS). This represents the proportion of phytochrome in the active Pfr form under a given spectrum.
A high PSS corresponds to open, unshaded conditions. A low PSS signals shading, competition, or far-red enrichment. Importantly, this is not a binary system. Plants continuously integrate PSS as a gradient signal and adjust growth accordingly.

This is the first key shift in perspective:
you are not just delivering photons—you are setting the phytochrome equilibrium.
The Red:Far-Red Ratio as a canopy intelligence signal

In natural sunlight, the red:far-red ratio typically sits around 1.1–1.2. Inside a plant canopy, however, this ratio collapses. Chlorophyll absorbs red light efficiently, while far-red wavelengths are transmitted and reflected deeper into the crop. In dense canopies, the ratio can drop to 0.1 or lower.
For the plant, this is a highly reliable signal of competition.
The response is not subtle. The plant reprograms its growth strategy toward rapid vertical expansion, reallocating resources to outgrow neighboring plants. This includes elongation of stems, changes in leaf morphology, and adjustments in developmental timing.
In controlled environments, this mechanism becomes a powerful design variable.
Even without dedicated far-red emitters, dense planting itself creates a far-red-enriched microclimate within the canopy. Conversely, a spectrum with minimal far-red can lock the crop into a permanently “open field” signal, regardless of plant density.
This is why far-red cannot be treated as a simple additive component.
It is a context-setting signal that defines how the plant interprets its entire environment.
Far-Red in grow lights and Photosynthesis: extending beyond PAR
The conventional definition of Photosynthetically Active Radiation excludes far-red light. From a strict single-photon efficiency perspective, this makes sense. Far-red photons alone contribute relatively little to carbon fixation.
However, this boundary does not fully capture how photosynthesis operates in real systems.
When far-red light is combined with shorter wavelengths, it enhances overall photosynthetic efficiency by preferentially exciting Photosystem I, complementing the activity of Photosystem II driven by red light. This interaction—often described as the Emerson Enhancement Effect—results in increased electron transport rates and improved CO₂ assimilation.

Experimental work, including studies by Park and Runkle, has shown that supplementing red/blue LED systems with far-red can accelerate plant growth and increase biomass accumulation under certain conditions.
The implication is practical and important:
light metrics based purely on PAR can underestimate the functional contribution of far-red-inclusive spectra.
At the same time, the response is not universally positive. Some studies indicate that far-red-driven growth increases fresh weight while reducing concentrations of compounds such as vitamin C and antioxidants. The outcome is species- and context-dependent, reinforcing the need for crop-specific spectral strategies.
Photoperiod control: manipulating time through spectrum

Beyond morphology and photosynthesis, the phytochrome system plays a central role in how plants measure time.
During darkness, the active Pfr form gradually reverts to Pr. This decay acts as a biological clock, allowing plants to estimate night length and regulate flowering accordingly.
Far-red light accelerates this conversion. By applying far-red at specific times—particularly at the end of the photoperiod—growers can manipulate the perceived duration of night without changing total light hours.
This enables precise control over flowering responses in both short-day and long-day crops. Techniques such as night-break lighting and end-of-day far-red treatments are widely used to influence flowering initiation, stem elongation, and developmental balance.
In crops like strawberry, controlled far-red exposure has been shown to shift the balance between vegetative propagation and reproductive development—demonstrating how spectral signals can directly influence commercial outcomes.
Canopy-Level effects: penetration and spectral uniformity for far-red grow light

Far-red light behaves differently from shorter wavelengths when interacting with plant tissue. While blue and red photons are absorbed near the leaf surface, far-red penetrates deeper into the canopy.
This has two system-level consequences. First, it improves activation of lower canopy leaves, contributing to more uniform photosynthesis across the plant. Second, it interacts with the plant’s own optical properties—reflected and transmitted light—to shape the internal spectral environment.
In multi-layer or high-density systems, this becomes critical. The spectrum experienced by a plant is not identical to the spectrum emitted by the fixture. It is modified by canopy structure, plant spacing, and optical design.
This is where fixture engineering matters. The spatial distribution of far-red emitters, combined with optical components from manufacturers such as LEDiL or Carclo, determines whether the red:far-red ratio is uniform across the crop—or varies significantly from plant to plant.
Uniform PPFD alone is not sufficient.
Spectral uniformity at canopy level is equally important.
From Spectrum to Strategy: engineering for phytochrome response
The inclusion of far-red light in modern LED systems is no longer incidental. It is increasingly intentional, reflecting a shift toward designing for photoreceptor response—not just chlorophyll absorption.
A clear example is the development of phytochrome-aware LED platforms such as Nichia’s Hortisolis™, which integrates far-red alongside blue and red channels to target multiple photoreceptor systems simultaneously. The goal is not simply broader spectrum coverage, but controlled activation of biological pathways that influence growth and yield.
At Lumistrips, this philosophy is applied through custom LED module design. By combining LEDs from leading manufacturers such as Nichia, Cree, Seoul Semiconductor, and OSRAM with application-specific optics and thermally optimized PCB platforms, it becomes possible to engineer spectra that directly target phytochrome-mediated responses.
This enables growers to move beyond static recipes toward dynamic strategies—adjusting spectral composition based on crop type, growth stage, and production objectives.
Far-Red light and photosystem: beyond phytochrome
Far-red light is typically discussed in the context of phytochrome signaling, but it also plays a direct role in photosynthesis through its interaction with Photosystem I (PSI).
The reaction center of PSI, known as P700, is tuned to absorb longer wavelengths, including far-red light around 700–730 nm. When excited, PSI boosts electrons to a higher energy state, enabling the production of NADPH. However, because PSII is less responsive to far-red light, relying on far-red alone can create an imbalance in the electron transport chain.
This is where the interaction between red and far-red becomes critical. Red light efficiently drives PSII, while far-red enhances PSI activity. When both are present, the two photosystems operate in synchrony, increasing overall photosynthetic efficiency. This principle underlies the Emerson Enhancement Effect and explains why supplemental far-red can improve plant performance when used in combination with red light.
What Happens when you change the Spectrum at different growth stages
The general recommendation to supplement far-red for growth is correct as far as it goes, but it flattens a more operationally important question: when in the crop cycle should far-red be present, and what happens if the spectral programme changes between early growth and the pre-harvest phase?
A 2024 study by Ju et al. from South China Agricultural University (Agronomy, 14, 55) addressed this directly using lettuce (Lactuca sativa L. 'Green Butter') in a hydroponic plant factory over a 35-day post-transplant production cycle.

The experiment compared four treatments against a white LED baseline of 250 µmol·m⁻²·s⁻¹ on a 16/8 h photoperiod.
The white-only control (W) maintained a stable R:FR of 12.36 throughout. These ratios bracket a wide range of phytochrome states, and the results show that plants tracked each shift in real time rather than retaining any morphological memory of the previous phase.
The first treatment, designated A, supplemented both red (660 nm) and far-red (730 nm) continuously throughout the entire growing period, bringing the R:FR ratio to 2.17.
The second, FRR, used far-red supplementation in the first ten days — lowering R:FR to 1.44 — then switched to red supplementation in the final ten days, raising R:FR to 14.22.
The third, RFR, did the reverse: red first at R:FR 14.22, then far-red at R:FR 1.44.
Four strategies, four Outcomes
Continuous supplementation of both red and far-red throughout the entire cycle (treatment A) produced the largest, heaviest plants in the trial. Total fresh weight increased by 53.76% compared with the white control, leaf area expanded significantly, and soluble sugar content rose by 39.98%. Vitamin E increased by 34.21%. These are compelling numbers for a grower whose primary objective is maximum harvestable biomass and who sells into markets where sugar content and vitamin E matter. The cost is a meaningful reduction in secondary metabolites: total flavonoid content fell by 55.95% and ferric-reducing antioxidant power declined by 35.41%. The mechanistic explanation is straightforward — plants under sustained low R:FR allocate metabolic resources toward structural growth and biomass rather than secondary metabolite synthesis. Polyphenols and flavonoids are energetically expensive to produce, and when the phytochrome system is continuously signalling shade, the plant deprioritises them.
The FRR sequence — far-red in the early vegetative phase, red in the final phase before harvest — represents the most balanced outcome across the full range of metrics assessed. Fresh weight increased by 26.10%, soluble sugars by 35.24%, and nitrate content fell by 31.52%. Importantly, the majority of nutritional indices remained comparable to the white control rather than declining. The authors attribute this to the late-stage shift toward high R:FR: as the plant's phytochrome state rises in the final days before harvest, the metabolic suppression associated with prolonged far-red exposure is partially reversed and secondary metabolite synthesis recovers.
From a commercial perspective this scheme delivers meaningful biomass gains alongside reduced nitrate — a quality marker of particular importance for fresh lettuce — without the antioxidant losses that continuous far-red produces. The authors explicitly recommend FRR as the preferred scheme for PFAL lettuce production when energy efficiency and nutritional quality are both priorities.
The RFR sequence produces a fundamentally different plant. Starting with red at R:FR 14.22 in the early phase establishes a flat, compact canopy consistent with high-PSS morphology. When the spectrum switches to far-red in the final phase and R:FR drops to 1.44, shade avoidance responses activate and the plant adopts a progressively more upright, narrow architecture. The result at harvest is a more vertical plant with a smaller footprint — an advantage in multi-tier vertical farms where canopy width constrains planting density. Individual plant fresh weight is lower than under treatment A or FRR, but the upright architecture can allow enough additional plants per unit area to recover or improve yield per square metre. Vitamin E content increased by 23.87%, but the late far-red phase also suppressed phenolics by 21.75%, flavonoids by 59.03%, and DPPH antioxidant activity by 19.02%. The sequence matters: finishing under far-red appears to suppress secondary metabolite synthesis regardless of what spectral regime preceded it.
The white-only control set the floor for yield across all treatments, with smaller plants and lower fresh weight than any supplemented scheme. It did, however, retain the highest antioxidant and flavonoid profile of all four conditions. For operations producing lettuce into markets where antioxidant content is a primary quality claim, white-only production with no spectral supplementation may be a defensible baseline — but it also illustrates precisely the headroom that well-designed spectral programming leaves on the table.
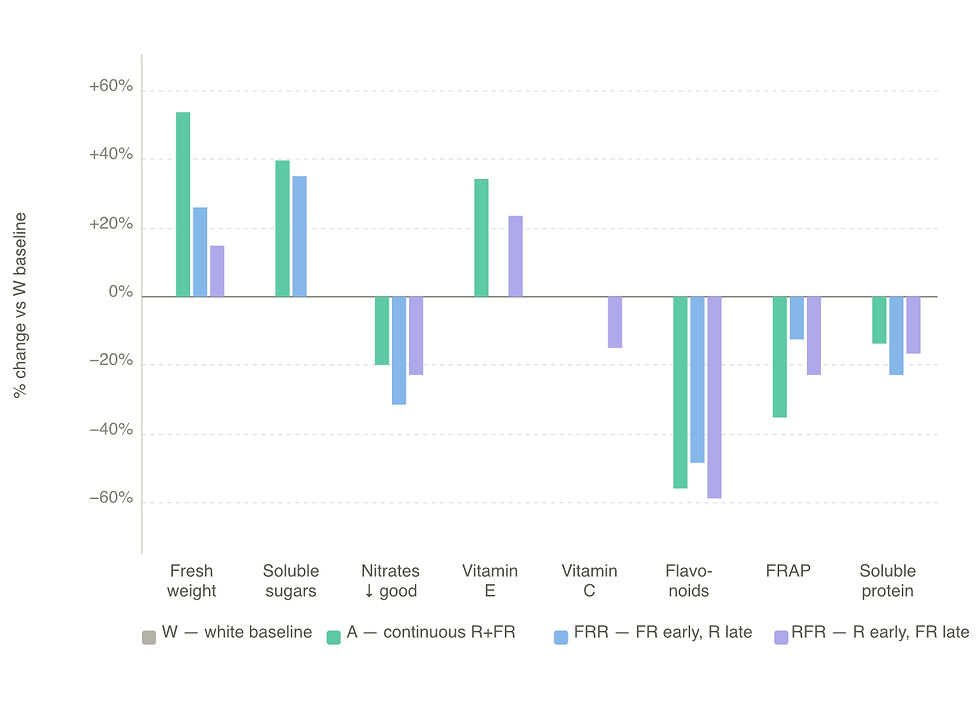
The study does not produce a single winner, because the four treatments pull in different directions across the metrics that matter commercially. Treatment A is the clear leader for growers selling by weight, delivering the strongest biomass and sugar numbers in the trial. FRR is the best all-rounder: it produces meaningful yield gains, preserves most nutritional indices, and cuts nitrate — which is why the authors recommend it as the default scheme for PFAL lettuce when both productivity and quality are in scope. RFR trades nutritional depth for architectural control, producing the most upright plants in the trial and a genuine density advantage in constrained growing systems, at the cost of the steepest antioxidant reductions of any supplemented treatment — flavonoids down 59.03%, FRAP down 22.86%, driven specifically by the low R:FR phase occurring right at harvest. Treatment W is the floor on yield and the ceiling on antioxidants. The practical takeaway is that the right spectral scheme is a market question as much as an agronomic one, and no fixed-spectrum system can serve all four outcomes simultaneously.
Summary of the study results:
Treatment | Results | Best For |
A (continuous R + FR throughout) | Maximum biomass and fresh weight: +53.76% total fresh weight, +39.98% soluble sugars, +34.21% vitamin E versus the white-only control. | Maximum yield |
FRR (far-red early, red pre-harvest) | Balanced yield-plus-quality outcome: +26.10% fresh weight and +35.24% soluble sugars while keeping most nutritional indices stable and cutting nitrate content by 31.52%. . From an energy-efficiency standpoint this recommend this scheme for PFAL lettuce production. | Best overall balance |
RFR (red early, far-red late) | The most upright, compact architecture. This doesn't maximise individual plant weight, but it can increase plants per square metre in multi-tier systems, recovering yield per unit area by a different route. It also raises vitamin E by 23.87%. | Morphology / density |
W (White only) | The worst performer against a combined yield-and-quality benchmark. However, W retained the best antioxidant and flavonoid profile, which means it is the least-worst option if phenolic content and FRAP are the primary quality target. | Best antioxidant and flavonoid profile |
Sequence Is a Design variable
The study's most important implication for lighting system design is that spectral sequence across growth stages is a lever that fixed-spectrum fixtures cannot pull. A lamp with a fixed R:FR ratio delivers one developmental trajectory regardless of how useful it would be to shift that ratio at transplant stage, mid-growth, or pre-harvest. The differences between FRR and RFR outcomes — plants that are similar in many growth metrics but structurally and nutritionally distinct at harvest — were produced entirely by swapping the order of the same two spectral conditions across the same 20-day window. That is not an argument for complexity for its own sake; it is an argument that the optimisation space available to a grower with a tunable multi-channel system is categorically larger than what is available with a fixed one. The right spectral strategy depends on what the market demands: maximum weight, minimum nitrate, upright architecture for density, or antioxidant preservation. With a fixed lamp, the grower picks one and accepts the trade-offs. With a programmable system, they can navigate between them.
Lumistrips: Built for Spectral Precision
This is exactly the design territory where Lumistrips' approach to custom LED module manufacturing is directly relevant. A production-grade horticulture module requires not just the right LEDs but the engineering infrastructure to deliver spectral control, thermal stability, and optical uniformity at scale.
Implementing these strategies in practice requires more than simply adding far-red LEDs to a fixture. It requires control—both spectral and temporal. Multi-channel LED systems, where red and far-red can be independently adjusted, enable growers to apply stage-specific light recipes without changing hardware.

This is where custom LED module design becomes critical. At Lumistrips, we develop horticulture lighting solutions that integrate high-performance LEDs from leading manufacturers such as Nichia, Cree, and OSRAM, combined with application-specific optics and thermally optimized PCB platforms. This allows precise control over spectral composition, uniformity, and intensity—ensuring that the intended phytochrome response is delivered consistently across the canopy.
More importantly, it enables the transition from static lighting setups to adaptive systems—where spectrum evolves alongside the crop. In this context, far-red is no longer an optional addition, but a key parameter in shaping plant development, yield, and quality.
FAQ – Far-Red Light and the Phytochrome System
What is far-red light in horticulture?
Far-red light refers to wavelengths between approximately 700 and 750 nm, just beyond the traditional Photosynthetically Active Radiation range. While less efficient for direct photosynthesis, it plays a critical role in regulating plant growth, morphology, and flowering through the phytochrome system.
What does the phytochrome system do in plants?
The phytochrome system is a photoreceptor mechanism that allows plants to detect the ratio of red to far-red light in their environment. It regulates key processes such as stem elongation, leaf expansion, flowering, and shade avoidance by switching between inactive (Pr) and active (Pfr) forms.
How does the red to far-red ratio affect plant growth?
The red:far-red ratio acts as a signal of competition. A high ratio (more red light) promotes compact growth, while a low ratio (more far-red light) triggers elongation and canopy expansion. This response allows plants to adapt to shading conditions and can be controlled in indoor farming.
Does far-red light contribute to photosynthesis?
On its own, far-red light has limited photosynthetic efficiency. However, when combined with shorter wavelengths, it enhances Photosynthesis through the Emerson Enhancement Effect, improving overall energy conversion and plant growth.
What is the Emerson Enhancement Effect?
The Emerson Enhancement Effect describes the synergistic interaction between red and far-red light. When both wavelengths are present, they activate Photosystem II and Photosystem I simultaneously, increasing the overall efficiency of photosynthesis beyond what either wavelength can achieve alone.
What is Phytochrome Stationary State (PSS)?
Phytochrome Stationary State (PSS) is a measure of the proportion of phytochrome in its active form under a given spectrum. It depends on the red:far-red ratio and determines how plants interpret their light environment, influencing growth patterns and development.
When should far-red light be used in plant growth?
Far-red light is most effective when applied strategically at specific growth stages. Early-stage far-red promotes canopy expansion, while late-stage far-red can influence plant architecture and flowering. Dynamic use across the crop cycle typically delivers better results than constant exposure.
What is the best red and far-red lighting strategy?
There is no single optimal strategy. Continuous red and far-red light maximizes biomass, while stage-based approaches—such as early far-red followed by red—offer a better balance between yield and quality. The ideal approach depends on crop type and production goals.
Does far-red light affect plant quality and nutrients?
Yes. Far-red light can increase biomass and sugar content, but it may reduce certain secondary metabolites such as antioxidants. Proper spectral balance is required to optimize both yield and nutritional quality.
Why is far-red important in vertical farming?
In vertical farming, far-red light improves canopy penetration and influences plant architecture, allowing better light distribution and higher planting density. This leads to more efficient use of space and increased yield per square meter.
Can LED grow lights be optimized for phytochrome control?
Yes. Modern LED systems can include independently controlled red and far-red channels, allowing precise adjustment of the spectral balance. This enables growers to influence phytochrome responses and tailor light recipes to specific crops and growth stages.
How do custom LED modules improve horticulture lighting?
Custom LED modules allow precise control over spectrum, intensity, and uniformity. By integrating red and far-red LEDs with optimized optics and thermal design, systems can be tailored to maximize plant performance, energy efficiency, and crop-specific outcomes.



Comments